
Teeth and vertebra - The fossils of megalodon
The teeth are by far the most common remains of C. megalodon with the larger teeth approaching 18 centimetres in height.
It is uncertain for how long people have been collecting C. megalodon teeth, but it was not until 1667 that science recognised them as shark teeth.
Before this time people generally and honestly thought that C. megalodon teeth were actually the tongues of dragons that had been petrified (turned to stone).
The truth about them came about when the Danish naturalist Nicolas Steno correctly identified one of these ‘dragon tongues’ and mentioned it in his book The Head of a Shark Dissected.
C. megalodon teeth are usually compared with those of the great white shark because of their superficial similarities of being triangular and serrated.
Because of this similarity, particularly with the serrations, it is thought that C. megalodon had the same biting style as the great white shark.
This would involve biting down upon its prey and then shaking its head from side to side so that the teeth serrations sawed through the flesh.
There have even been what are close to being complete sets of C. megalodon teeth from the same individual found together.
These reveal that C. megalodon was like other sharks in that the largest teeth were at the front of the mouth.
Also the further towards the front the teeth were, the more evenly triangular they became.
The more sideward the teeth went towards the rear of the jaws, the more inwardly curved they became.
These teeth would hook into the flesh of the prey making it harder for it to escape while the front teeth did the cutting.
One of the most common myths most often told about C. megalodon is that it is only known from teeth.
In reality the teeth are by far the most common fossils of C. megalodon, but a small and growing number of vertebras have also been recovered.
In shark biology in general, the skeleton is always made up of ‘soft’ cartilage, but the vertebrate are made up of harder calcified cartilage.
This means that while the vertebra can and often do decompose, they can survive for longer increasing the chance of them getting fossilised.
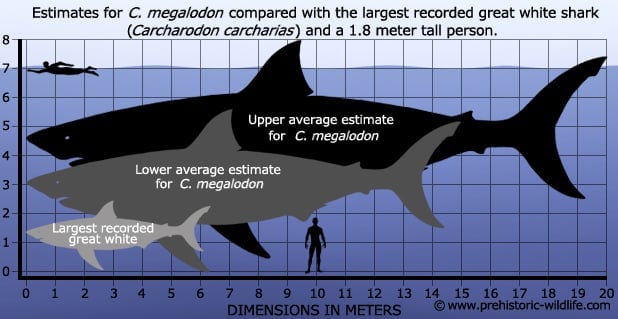
Size -Estimating the upper size of megalodon
Ever since the teeth of C. megalodon were realised to be the teeth of sharks most people immediately asked, ‘but how big was the shark?’.
The honest answer is that we don’t exactly know for certain because there is no complete specimen that someone can take a tape measure to.
So instead palaeontologists and ichthyologists have to work with what they have, in this case the teeth.
They have been many studies into estimating the length of a shark based upon analysing teeth, the four most commonly referred to being here.
One method once used to gauge the length of C. megalodon was from measuring the height of the tooth enamel.
This method was developed by John E. Randall in 1973 and resulted in an estimate of 13 meters.
However the method is not considered accurate by the wider scientific community as the method was first originated by comparing the teeth of the great white shark, and although these teeth appear similar at a glance, they are actually quite different from the teeth of C. megalodon.
Also the amount of enamel on a tooth can vary even amongst teeth of the same species through wear and poor preservation.
Another method of estimation developed in 1996 by Michael D. Gottfried, Leonard J. V. Compagno and S.
Curtis Bowman works on the basis of establishing a correlation between the slant height of a tooth and the length of the shark.
Slant height is not the total height but the length of the tooth tip to the lateral edge, and is the actual cutting length of the tooth.
Again this study was based upon the study of great white teeth, and this yielded an estimate of 15.9 meters.
However even if this method is reliable, it can only give estimates from the given teeth, and not necessarily the species.
In 2002 Dr. Clifford Jeremiah established another method based upon the width of a tooth root, the bit below the crown that is covered by flesh.
This is a simple principle that the width of the tooth crown can be used to figure out the width of the jaw, which in turn can be used to work out the length of the shark.
In the simplest terms it works on the basis that there is 1 centimetre of tooth root for roughly every 129 centimetres of body length.
This resulted in a 15.5 meter length estimate for a tooth which had around 12 centimetres of root.
Also in 2002, Dr. Kenshu Shimada developed another more complex method based upon the proportions of the tooth crown.
When this method was used upon the tooth that had been used by Gottfried et al. in 1996, the estimated length came back as 15.1 meters.
The maximum estimate ever yielded from this method was obtained from a particularly large tooth from Panama which gave a body length of 16.8 meters.
Going with an estimated body length in the range of 15-16 meters is considered quite safe for C. megalodon, with higher estimates approaching 17 meters being considered realistically possible.
Larger estimates do exist with claims that C. megalodon could approach and even exceed lengths of 20 meters, however most of these estimates are considered to be only possibilities and do not reflect any current fossil evidence.
Still, even at a more modest 15 meters, C. megalodon would still have dwarfed any other shark either alive or extinct with even a Great White shark paling in comparison to its gigantic size.
How much C. megalodon weighed is another matter of study, although its one than can produce an even wider range of estimates.
Current estimates are once again based upon comparison to the great white shark, with an estimate for a 15.9 meter C. megalodon being gauged at 47 metric tons.
Larger estimates include 59 metric tons for a 17 meter C. megalodon and 103 metric tons for a 20.3 meter C. megalodon.
As you may have noticed, the weight of a C. megalodon does not increase by a constant amount with length, which is why a 20.3 meter shark is roughly a third longer than a 15.9 meter shark, yet weighs more than double the slightly smaller specimen.
This is because you have to realise that the shark is not just longer, it is also proportionately thicker and bulkier with increased muscle mass to move the larger frame.
This also means that an animal, be it a shark or any other kind, will always reach a size from which it can grow no further because of what its habitat can support in terms of food, a natural fact based upon the logic that a bigger body needs a greater intake of food for energy to fuel it.
Possibly Biology - The inner workings of megalodon
Because of its large body size C. megalodon is thought to have almost certainly have lived with the effects of gigantothermy.
This is where a creature is so massive that its body holds onto heat with the outer layers of muscle and flesh actually insulating the internal organs from the environment.
This causes an increase in metabolism which in turn makes the creature more active. C. megalodon may have taken the process of gigantothermy even further by directing the flow of warmer blood into its head and snout like the great white shark is known to do.
This would have the effect of making the brain and sensory organs of C. megalodon such as smell, sight and electroreceptive ampullae operate at warm blooded metabolism levels.
The fact that C. megalodon teeth are known from every continent is proof that it had a cosmopolitan distribution like many other successful marine predators.
However the distribution of C. megalodon seems to have become reduced as the oceans cooled, especially towards the end of the Pliocene.
The overall appearance of C. megalodon would most likely have been very similar to the lamniform sharks that are swimming in the ocean today, although its large size and inherent power means that it was probably quite stocky in comparison to modern sharks.
Also the cartilaginous skeleton would have likely been more robust to deal with the greater stresses of larger muscles and more powerful prey items.
C. megalodon probably had a movable upper jaw that not only moved up and down, but could be projected forwards independently from the rest of the body.
This kind of jaw can be seen in modern sharks today and is why a shark that is about to bite down on something looks very different from a shark that is just cruising.
When the upper jaw is projected forward, the upper teeth and jaw actually become visible, whereas they are normally hidden by the mouth.
With a jaw that moves in this way, C. megalodon would latch onto its prey with its lower jaw holding it in place and then shake its head from side to side so that the teeth serrations sliced off a chunk of flesh.
The upper jaw is then retracted which has the effect of pulling the chunk of flesh into its mouth. The shark then repeats this process until it has eaten its fill.
An area of study immediately associated with the discovery of the teeth is the estimation of the bite force that C. megalodon was capable of.
This is an important thing to know as how strongly an animal can bite down upon something can lead to clues to what kind of prey an animal ate to how it ate them.
In 2008 a biomechanical computer model was created to estimate the bite force of great white sharks.
When this model was applied to C. megalodon it was revealed that a 15.9 meter C. megalodon, a large average, was capable of exerting a bite force of 108, 514 newtons, just over 11 metric tons.
When the bite force for a 20.3 meter C. megalodon, the upper end of the estimated potential C. megalodon size scale, was estimated the result was 182, 201 newtons, over 18.5 metric tons.
This means that even the lower estimates for C. megalodon still give it a much higher bite force than the ferocious placoderm Dunkleosteus, and even higher than the mighty Tyrannosaurus rex.
Another thing to be considered is the fact that C. megalodon probably did shake its head from side to side like other sharks with a similar dentition.
This meant that the actual forces subjected to the unfortunate prey of C. megalodon were likely even greater.
Whereas most of the study for C. megalodon centres on the teeth, one area that is often overlooked are the vertebrae.
Because of the small number of known vertebrae not everyone is able to study them like they can the teeth, but one thing that the vertebra clearly show are concentric rings.
These concentric rings are essentially the same as the growth rings that you can see in a tree stump, and they are also visible in other modern sharks that are swimming the oceans today.
Counting these concentric rings has yielded estimates of 25 to 40 years of age for the C. megalodon known from fossils, with palaeontologists suggesting that C. megalodon may have been able to live for even longer than this.
Young megalodon - Pups and Nursery grounds
Given their large size and pelagic lifestyles, C. megalodon are thought to have given birth to live young.
Exactly how C. megalodon did this though is still open to debate as sharks have two methods of doing this.
The first is termed viviparity, and this is where the pup, which is what a baby shark is called, grows inside the mother until it is ready to be born.
Shark pups born in this method are fed nutrients via an umbilical cord, and once the pup is born, the placenta is usually ejected straight afterwards.
The second method is ovoviviparity, and this is where the pup develops inside an egg.
However in sharks that display ovoviviparity the egg is not laid but retained inside the mother shark.
Pups of these sharks are not attached to their mother by an umbilical cord, but instead use up the egg yolk in the course of their development.
In both types of development the pups are conscious and able to swim under their own power when they pass out of their mother’s body.
C. megalodon pups are usually depicted as coming out tail first and while this is deemed the usual method, some shark species today are actually born head first.
Also like we see in sharks today, C. megalodon probably didn’t give birth to their pups just anywhere, but instead chose what are called nursery areas.
A nursery area is where a young shark can live and hunt while being safe from other larger predators.
Sharks in nursery areas are considered generalists that attack and eat all manner of creatures including fish, cephalopods (like octopus and cuttlefish), turtles, and pretty much anything else they can catch.
To identify potential nursery areas palaeontologist look for concentrations of smaller C. megalodon teeth.
One area that seems to be plentiful in these teeth is central America and the most Southern areas of the United States.
Going back towards the Oligocene, Panama did not exist because the higher ocean levels of the time submerged much of the area.
This area was known as the Central American Seaway, and formed an oceanic pass between the Pacific and Atlantic oceans.
The area likely had a huge expanse of shallow waters that were simply not deep enough for larger predators to operate in, making them relatively safe for the much smaller C. megalodon pups.

Fossil evidence strongly suggests that during the Miocene the shallow waters between North and South America had a very high population of juvenile C. megalodon, while the larger adults roamed the deeper waters of the open ocean (red).
Fossil deposits also indicate that early cetaceans such as whales (purple) used what was then known as the Central American Seaway as a passage between Pacific and Atlantic waters.
High volcanic activity as well as falling sea levels closed this passage off resulting in a reduction of available prey as well as a dramatic shift in ocean currents. Above right is Central America as we know it today, with the warm water ocean currents.
Study of the smaller teeth indicates that the C. megalodon in the nursery areas were as small as 2 to 3 meters long.
However this does not mean that they were this large when they were first born, just that the teeth came from juvenile individuals that were active in the area.
As the young C. megalodon grew older they would also grow bigger, and as such they would eventually have to leave the shallows for open ocean life.
This would begin the second stage of their lives where they would have to specialise in attacking large ocean going creatures.
Prey items - What did megalodon eat?
One thing that needs to be immediately cleared up is that C. megalodon did not eat dinosaurs.
This is a myth spread around in popular culture, particularly in films and novels where the C. megalodon ‘villian’ gets built up to be more dramatic.
C. megalodon does not enter the fossil record until the late Oligocene, about 36 million years after the dinosaurs went extinct at the end of the Cretaceous making it impossible for C. megalodon to have eaten dinosaurs (for a shark that really could have fed upon dinosaurs and large marine reptiles look up Cretoxyrhina).
The preferred food for C. megalodon seems to have been cetaceans and particularly small to medium sized whales.
There is also evidence that C. megalodon attacked and ate large sea turtles which were presumably too slow to escape with even their shells not providing protection against the colossal bite force of C. megalodon.
However what C. megalodon hunted depended upon the age of the individual with smaller C. megalodon hunting animals like dugong, and larger older C. megalodon hunting the larger whales.
The attack strategy of C. megalodon was not exactly refined, but it was effective at taking down whales.
The vertebrae of some cetaceans display compression damage which has been interpreted to have been caused by a sudden and massive impact from below.
This allowed for the reconstruction of a scenario where C. megalodon would approach a whale from below so as to avoid being seen by its target.
Once it had lined itself up for a strike, C. megalodon would use its powerful muscles to propel itself to the surface at high speed and slam into the whale from underneath.
If the whale did not end up in the jaws of C. megalodon it would have very likely been stunned by the impact, allowing C. megalodon time for a killing bite.
However at least one fossil vertebra is known to exist that shows it was subjected to this style of attack, yet still managed to heal.
This shows that in this case the lucky animal did not just survive the attack, it also lived long enough for the injuries to heal.

Examination of fossils that seem to have come from C. megalodon prey show that C. megalodon actually targeted the bony areas like the ribcage.
Here C. megalodon had two things in its favour, extremely robust teeth that did not break easily, and a crushing bite force that could easily break bones which in turn caused large scale injuries to the internal organs that they were supposed to protect.
Further support for this method comes from compression fractures of the teeth where they have been blunted, which suggests strong impacts with a hard substance such as bone.
When attacking larger whales that were feasibly too big for a bite to an area like the ribs, C. megalodon changed its tactics. Instead of going for the critical organs, C. megalodon attacked the tail to try and immobilise its prey.
This is a very intelligent strategy, as even though sharks are almost constantly swimming forward so that they can breathe, they can only maintain extremely fast pursuit speeds for short durations.
This is because in sharks the white muscle (about 90% of the total muscle mass) which is used to provide sudden bursts of speed gets tired very quickly, whereas a sharks red muscle (roughly 10% of the total muscle) has less power, but an incredible amount of endurance which is why the lesser red muscle is used for standard cruising.
By crippling a large whale, a C. megalodon could take its time feeding instead of over exerting itself.
Some people have made claims that C. megalodon was simply too large to hunt and could only have been a scavenger.
In the face of overwhelming fossil evidence that shows injuries, not just tooth marks, to many large cetaceans, such a claim is not just considered unlikely but almost impossible.
While most sharks, and carnivorous animals in general, will take the opportunity to feed from a carcass, doing so does not make them exclusive scavengers.
Also marine animals that live by just scavenging tend to be bottom feeders that wait for dead animals to sink to the bottom of the sea. Estimating the size of C. megalodon has also brought estimates of how much food it would take to keep it going.
Amounts vary greatly but range between 600 to 1200kg of food every day.
This is a tremendous amount of food for a scavenger as scavengers tend to be adapted to require very little energy expenditure because they don’t know when or where their next meal is coming from.
When taking all of the fossil evidence, biometric models and knowledge about shark lifestyles and biology into account, the result is that scavenger is the least likely method of survival for C. megalodon.

Extinction - Why did megalodon disappear?
C. megalodon disappears from the fossil record near the end of the first stage (Gelasian) of the of the Pleistocene 1.8 million years ago.
This disappearance is marked by the steady decline in C. megalodon fossils until they disappear completely.
While there are a few theories as to why C. megalodon went extinct it seems most likely that a sequence of changing events brought about its downfall rather than just one thing.
The trigger event for the extinction of C. megalodon seems to have been global cooling.
To start, if C. megalodon had a warm blooded metabolism through gigantothermy, then it would need a higher calorie intake than an entirely cold blooded creature of similar size.
The colder the water the greater the difference, meaning that C. megalodon would have needed an even greater amount of food to deal with the temperature reduction.
Additionally gigantothermy is still no substitute for a true warm blooded metabolism and a shark in cooler waters would still have possibly been more sluggish than it would have been in warmer waters, something that would further impede its ability to hunt.

A knock on effect of colder global temperatures is that large amounts of water began to solidify into ice as evidenced by the presence of vast ice sheets across the Northern Hemisphere.
The presence of more ice meant that the sea level dropped and the most dramatic consequence of this was the creation of the Isthmus of Panama, something that was also helped by new land formations being built up by ongoing volcanic activity in this area.
This essentially created a land bridge between North and South America as well as isolating the Pacific and Atlantic oceans from each other at this point.
The immediate result of the creation of the Isthmus of Panama was the closure of the Central American Seaway which seemed to have been used as a key migration route for whales, as evidenced by the large concentration of whale fossils.
This coincides with an overall decline in whale species with much less than half of the Pleistocene whales surviving to the present era. Today there are only 6 genera of whales as opposed to over 20 genera during the Miocene.
The remaining whales were still migratory but seemed to have preferred Polar Regions, presumably for the greater abundance of invertebrate food that Baleen whales are adapted to eat.
The toothed whales do not seem to have been a viable option either as their numbers were also dramatically reduced with the sperm whale being the only large toothed whale to survive to today.
With C. megalodon restricted to the warmer ocean waters it no longer had constant year round access to the food supply that it was most adapted to kill.
The huge size of C. megalodon undoubtedly worked against it during these times as the only other prey available were smaller, faster, and even if caught did not provide the same level of sustenance as the larger whales did.
Cannibalism has also been suggested as a possible survival strategy for C. megalodon, but this would only work as long as there were other C. megalodon to eat.
If this did indeed happen, then all that cannibalism would do is thin out the C. megalodon numbers even further, which in turn would limit the numbers that would reproduce.
Linked to this is the potential loss of nursery areas caused by changing sea levels.
In fact the very creation of the Isthmus of Panama also seems to have removed one such Nursery area, as evidenced by large numbers of juvenile C. megalodon teeth from this area.
Another suspected nursery area was Maryland which was so far north the waters may have become too cold to support C. megalodon.
The loss of nursery areas means that the C. megalodon young would themselves have been more susceptible to predators, perhaps even other C. megalodon as they tried to survive.
The final theory involves the rise of new predators with special references made towards the evolution of raptorial delphinids, which today are represented by the Orca, also known as the killer whale.
As the Numbers of C. megalodon declined the numbers of delphinids increased.
However it is difficult to say if the rise of these new predators played a part in the decline of C. megalodon as it could equally be the decline of C. megalodon that allowed the new predators room to thrive.
There is fossil evidence that shows predator/prey interaction between C. megalodon and the delphinids as exhibited by C. megalodon tooth marks on delphinid bones.

Last survivors?
Some people think that C. megalodon survived the Pleistocene era and was still swimming the oceans as recently as the Holocene era.
Their proof for this claim comes from a partial C. megalodon tooth that was discovered in 1872 by the crew of HMS Challenger which when tested in 1959 was thought to be only 10,000 years old.
However this test measured the levels of manganese dioxide on the fossil, a method which is now considered flawed due to the varying degrees of manganese dioxide that can build up on different fossils, even from the same era.
When the tooth was submitted to later radio carbon dating techniques, the tooth was found to have too low a nitrogen level to allow for testing.
As such the tooth has since been deemed untestable and previous estimates of C. megalodon becoming extinct during the early Pleistocene remain valid.

Classification - Is megalodon related to the great white shark?
Perhaps the greatest point of controversy over C. megalodon is if it is actually related to the Great White Shark that we know today.
Comparisons between C. megalodon and the great white mostly come about from the principal that C. megalodon was the biggest shark, and the great white is the biggest shark that we know of today.
Also the teeth between the two are often seen as roughly similar. Supporters of C. megalodon within Carcharodon point to the tooth similarity as being the result of both C. megalodon and the great white being descended from Palaeocarcharodon orientalis.
The problem with comparing C. megalodon to the great white on the basis of similar teeth is that the only similarities that exist are that both sharks have teeth that are triangular and serrated.
Beyond this the teeth of the great white are more gracile, being much thinner than those of C. megalodon.
Also while C. megalodon is thought to have an overall similar tooth layout to the great white shark, the third anterior tooth (third front tooth from the centre of the upper jaw) of C. megalodon is different in that it points down like the first two, different to how it appears in the great white.
C. megalodon anterior teeth also have a characteristic ‘scar’ shaped like a chevron that lies between the crown and root of the tooth, something that is absent in the great white.
An alternative to placing C. megalodon within the Carcharodon shark genus would be to place it within the older Carcharocles genus.
The main argument for this placement is that another large ancient shark named Carcharocles auriculatus is thought to actually be an ancestor to C. megalodon.
With teeth that measured up to almost 12 centimetres in length Carcharocles auriculatus was big but roughly a third smaller than C. megalodon, if you scale it to an 18 centimetre C. megalodon tooth.
Even so, it is quite possible that Carcharocles auriculatus could have grown larger, giving rise to C. megalodon as predators in all environments have a tendency to keep growing bigger until their environment can no longer support further growth.
Such a placement of C. megalodon within Carcharocles would actually complete a transition where sharks lost lateral cusps to their teeth.
This transition begins with the lateral tooth cusps that are clearly present in Otodus obliquus, the reduced tooth cusps in Carcharocles auriculatus, to no cusps in C. megalodon.
These teeth also have chevron shaped scars where the crown meets the root, something which is absent from great white teeth.
Another theory suggests that C. megalodon was ancestral to the great white shark and that over time the shark simply grew smaller.
The main problem with this thinking however is that the great white shark was actually swimming in the ocean long before C. megalodon went extinct, with fossil teeth of the great white shark appearing back in the mid Miocene period 16 million years ago, over 14 million years before C. megalodon went extinct.
Supporters of the theory still insist that the great white could have evolved from a smaller species of C. megalodon.
Those familiar with the sabre toothed cat Smilodon may be aware that there were three quite different species of the same genus that not only seem to be descended from the same ancestor, but for a time were all active together in the same time period.
It is not too much of a stretch that similar occurrences could happen in other animal groups.
Still, there appears to be no transitional link that shows the change From C. megalodon into the great white as the teeth would not just grow smaller, they would steadily change into the great white form.
If indeed related, it is more likely that the great white shares a common immediate ancestor with C. megalodon.
In 2012 the theory that C. megalodon and Carcharodon carcharias are not related got a little more support with the description of a new species of Carcharodon, Carcharodon hubbelli, also known as Hubbell’s white shark.
The teeth of Carcharodon hubbelli have been interpreted by some to be transitional in form, linking Carcharodon carcharias with Isurus genus which houses the mako sharks.
Because most researchers do not consider there to be a direct link between mako sharks and megatoothed sharks like C. megalodon, this might suggest that Carcharodon carcharias is indeed separate from C. megalodon, and that by extension C. megalodon should be placed within the Carcharocles genus.
Any similarities in the overall morphology of C. megalodon and the great white are most likely the result of evolution rather than genetic breeding.
This basic body shape is called fusiform, or more loosely ‘torpedo shaped’, and is based upon a pointed front rising to a broad centre before tapering off to another rearward point.
This form has repeated itself in nature countless times, and is certainly not unique to just sharks as it is simply the most efficient form for submerged aquatic travel.
The megalodon shark species has also been regarded to some to belong to either the Procarcharodon or the Otodus genus.
This thinking is down to research that suggests a visible transition from the teeth of the Otodus type genus through to those of the megalodon species which lived later.
Again however, differences between researchers vary greatly as to which genus the megalodon species belongs to.
Further Reading
- – Recherches sur les poissons fossiles/par Louis Agassiz – Neuchatel :Petitpierre. p. 41. – Louis Agassiz – 1833-1843.
- – Size of the Great White Shark (Carcharodon) – Science Magazine 181 (4095): 169–170 – John Randall – 1973.
- – Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoi’n – Bulletin of the Geological Society of Denmark (Copenhagen: Geologisk Museum) 32: 1–32. – Svend Erik Bendix-Almgreen – 1983.
- – Catalogue of Cuban fossil Elasmobranchii (Paleocene to Pliocene) and paleogeographic implications of their Lower to Middle Miocene occurrence – Bolet�n de la Sociedad Jamaicana de Geolog�a (Cuba) 31: 7–21 – M. Iturralde-Vinent, G. Hubbel & R. Rojas – 1996.
- – The Megatooth shark, Carcharodon megalodon: Rough toothed, huge toothed – Mundo Marino Revista Internacional de Vida (non-refereed) (Marina) 5: 6–11. – J. C. Bruner – 1997.
- – Fossil sharks from Jamaica – Bulletin of the Mizunami Fossil Museum. pp. 211–215. – Stephen Donovan & Gunter Gavin – 2001.
- – An associated specimen of Carcharodon angustidens (Chondrichthyes, Lamnidae) from the Late Oligocene of New Zealand, with comments on Carcharodon interrelationships – Journal of Vertebrate Paleontology 21 (4): 730–739. – M. D. Gottfried & R. E. Fordyce – 2001.
- – The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae) – Journal of Fossil Research (Japan) 35 (2): 28–33. – Kenshu Shimada – 2002.
- – New Record of the Lamnid Shark Carcharodon megalodon from the Middle Miocene of Puerto Rico – Caribbean Journal of Science 39: 223–227. – Angel M. Nieves-Rivera, Maria Ruizyantin & Michael D. Gottfried – 2003.
- – The Miocene Climatic Optimum: evidence from ectothermic vertebrates of Central Europe – Palaeogeography, Palaeoclimatology, Palaeoecology 195 (3–4): 389–401 – M. B�hme – 2003.
- – Age of Carcharocles megalodon (Lamniformes: Otodontidae) : A review of the stratigraphic records – The Palaeontological Society of Japan (PSJ) (Japan) 75 (75): 7–15. – Hebe Hideo, Goto Mastatoshi & Kaneko Naotomo – 2004.
- – Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage – Caribbean Journal of Science 40 (3): 362–368. – O. Aguilera & E. R. D. Aguilera – 2004.
- – Tracing the ancestry of the Great White Shark – Journal of Vertebrate Paleontology 26 (4): 806–814 – K. G. Nyberg, C. N. Ciampaglio & G. A. Wray – 2006.
- – Late Neogene Oceanographic Change along Florida’s West Coast: Evidence and Mechanisms – The Journal of Geology (USA: The University of Chicago) 104 (2): 143–162. – Warren D. Allmon, Steven D. Emslie, Douglas S. Jones & Gary S. Morgan – 2006.
- – Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite? – Journal of Zoology 276 (4): 336–342. – S. Wroe, D. R. Huber, M. Lowry, C. McHenry, K. Moreno, P. Clausen, T. L. Ferrara, E. Cunningham, M. N. Dean & A. P. Summers – 2008.
- – Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles – Caribbean Journal of Science. 44 (3) pp. 279–286. – Roger Portell, Gordon Hubell, Stephen Donovan, Jeremy Green, David Harper & Ron Pickerill – 2008.
- – Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguan� Formation – Pal�ontologische Zeitschrift (Springer Berlin) 82 (2): 204–208. – Orangel A. Augilera, Luis Garc�a * Mario A. Cozzuol – 2008.
- – Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama – PLoS ONE (Panama: PLoS.org) 5 (5): e10552 – Catalina Pimiento, Dana J. Ehret, Bruce J. McFadden & Gordon Hubbell – 2010.
- – The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic – Revista Espa�ola de Paleontolog�a (Portugal) 25 (1): 1–6. – Miguel Telle Antunes, Ausenda C�ceres Balbino – 2010.
- – Patterns and ecosystem consequences of shark declines in the ocean – Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055–1071. – Francesco Ferretti, Boris Worm, Gregory L. Britten, Michael J. Heithaus & Heike K. Lotze – 2010.
- - Origin of the white shark Carcharodon (Lamniformes: Lamnidae) based on recalibration of the upper Neogene Pisco Formation of Peru - Palaeontology 55(6):1139-1153 - D. J. Ehret, B. J. MacFadden, D. S. Jones, T. J. DeVries, D. A. Foster & R. Salas-Gismondi – 2012.
- – Evolution of white and megatooth sharks, and evidence for early predation on seals, sirenians, and whales – Natural Science (Czech Republic) 5 (11): 1203–1218. – C. G. Diedrich – 2013.
- – Sharks and Rays (Chondrichthyes, Elasmobranchii) from the Late Miocene Gatun Formation of Panama – Journal of Paleontology 87 (5): 755–774 – Catalina Pimiento, Gerardo Gonz�lez-Barba, Dana J. Ehret, Austin J. W. Hendy, Bruce J. MacFadden & Carlos Jaramillo – 2013.
- – When Did Carcharocles megalodon Become Extinct? A New Analysis of the Fossil Record. – PLOS ONE. 9 (10): e111086. – C. Pimiento & C. F. Clements – 2014.
- – Body-size trends of the extinct giant shark Carcharocles megalodon: a deep-time perspective on marine apex predators. – Paleobiology. 41 (3): 479–490. – C. Pimiento & M. A. Balk – 2015.
- – Record of Carcharocles megalodon in the Eastern Guadalquivir Basin (Upper Miocene, South Spain). Estudios Geol�gicos. 71 (2): e032. – M. Reolid & J. M. Molina – 2015.
- – Geographical distribution patterns of Carcharocles megalodon over time reveal clues about extinction mechanisms. – Journal of Biogeography. 43 (8): 1645–1655. – C. Pimiento, B. J. MacFadden, C. F. Clements, S. Varela, C. Jaramillo, J. Velez-Juarbe & B. R. Silliman – 2016.
- – The size of the megatooth shark, Otodus megalodon (Lamniformes: Otodontidae), revisited. – Historical Biology: 1–8. – Kenshu Shimada – 2019.
- – The Early Pliocene extinction of the mega-toothed shark Otodus megalodon: a view from the eastern North Pacific. – PeerJ. 7: e6088. – R. W. Boessenecker, D. J. Ehret, D. J. Long, M. Churchill, E. Martin & S. J. Boessenecker – 2019.
- – The transition between Carcharocles chubutensis and Carcharocles megalodon (Otodontidae, Chondrichthyes): lateral cusplet loss through time. – Journal of Vertebrate Paleontology. 38 (6): e1546732. – V. J. Perez, S. J. Godfrey, B. W. Kent, R. E. Weems & J. R. Nance – 2019.
- – Body dimensions of the extinct giant shark Otodus megalodon: a 2D reconstruction. – Scientific Reports. 10 (14596): 14596. – J. A. Cooper, C. Pimiento, H. G. Ferr�n & M. J. Benton – 2020.








