In Depth
The holotype fossils of Altirhinus were originally classified as a species of Iguanodon, I. orientalis back in 1952. However Iguanodon has long suffered from the wastebasket taxon effect where remains of superficially similar dinosaurs have been attributed to the genus, in part due to its long history and seemingly wide distribution of fossils being attributed to the genus (in fact Iguanodon only just missed out on being the first known Dinosaur by a matter of months, Megalosaurus was the first to officially be named). New study the type specimens of I. orientalis in 1996 found that this species was actually no different from the main Iguanodon species I. bernissartensis that was based upon European material. Another discovery of the time however was that not all of the fossils previously assigned to I. orientalis were a match for Iguanodon, a discovery that led to the creation of Altirhinus in 1998 as a new home for these different fossils. The type species name of this genus is A. kurzanovi, and is in honour of the Russian palaeontologist Sergei Kurzanov.
The limbs of Altirhinus are interesting as with most iguanodonts the rear legs are longer and well developed for supporting the body weight, while the fore limbs have ‘hands’ that have tightly packed central digits with bony growths analogous to hooves. This indicates that Altirhinus was capable of both bipedal (two legged) and quadrupedal (four legged) postures, though it is uncertain how much time Altirhinus, and indeed its relatives, spent in either one. Some reconstructions portray iguanodonts as being mostly bipedal, but dropping to all fours to browse upon low growing vegetation. Other reconstructions have them as being mostly quadrupedal but balancing upon their back legs when reaching up to feed upon higher growing vegetation. Which one is more accurate would depend upon the actual behaviour of the living dinosaur, which at this point can only be guessed at. What is more certain is that Altirhinus and its relatives would have been able to run on two legs to try and escape predators, though how long they could maintain this kind of locomotion is again uncertain.
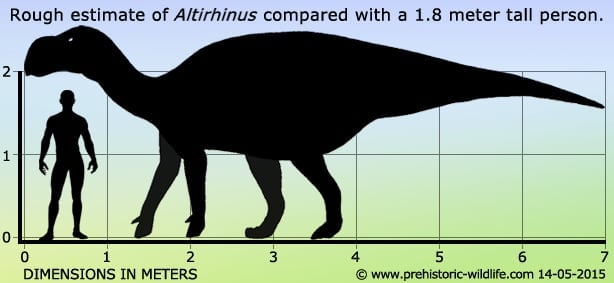
Altirhinus seems to have at least been a medium sized ornithopod that might have grown closer in size to the larger genera of the group. The exact size is uncertain for now because of the incomplete nature of available fossils which means that size estimates are based upon comparisons of more complete genera. This of course depends upon how much like Altirhinus was to the genera it is being compared to, and of course can only give us a rough idea as to the true size. Studies of other genera have led to the conclusion that Altirhinus should be placed within the Hadrosauroidea a group of dinosaurs descended from the Iguanodonts that first came into fruition back in the Jurassic. Dinosaurs within the Hadrosauroidea would eventually evolve into the hadrosaurids of the late Cretaceous. Among others, Altirhinus is thought to be more advanced than Fukuisaurus from Japan, but not less developed than Eolambia of the USA. Altirhinus might also be a relative to Muttaburrasaurus from Australia due to the striking similarity in the nasal arch of these two genera.
The high nasal arch seen in the skull of Altirhinus was the inspiration for the genus name which translates to English as ‘high snout’. Although uncertain due to lack of preservation, this arch might have existed for a special growth of soft tissue. If true, then this could have been for anything from visual/audial display, to some other unknown purpose. Another feature is the strong downward curve of the front portion of the mouth which might have been an adaptation to help the incisor teeth crop certain kinds of vegetation. Like with many other herbivorous animals, there was a toothless gap between the incisors and the batteries of molar teeth in the posterior portion of the mouth that is referred to as the diastema. The mouth would work by cropping plants with the front while also passing them back to be processed by the rear molars. It is also plausible that as a living dinosaur, Altirhinus also had cheeks covering the molar cheeks so that bits of plant would not fall out of the mouth while being processed between the teeth.
Further Reading
– On Asian ornithopods (Dinosauria: Ornithischia). 3. A new species of iguanodontid dinosaur. – Zoological Journal of the Linnean Society 122:291-348. – D. B. Norman – 1998.








