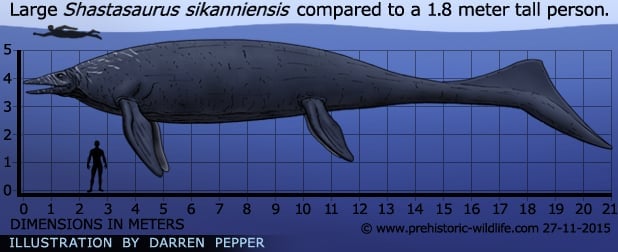
Shastasaurus species and fossil distribution
Shastasaurus hit the record books in 2011 with the re-examination of a set of ichthyosaur remains from British Columbia that were initially described as a new species of Shonisaurus back in 2004.
Study of the remains has found that while they are similar, the remains are a better match for Shastasaurus, and so the remains were moved from Shonisaurus to create a new third species, Shastasaurus sikanniensis.
This confusion is not surprising however when you consider that these two ichthyosaurs are thought to be very similar to each other in not just form but predatory behaviour as well.
Although the material referable to S. sikanniensis has allowed for a new upper size of the genus to be established, it is Chinese fossil material referable to the second species, S.liangae named in 2000 that has allowed for considerably more accurate reconstructions.
The type species of Shastasaurus was only ever known from fragmentary remains with additional fossils being attributed to it later. More complete S. liangae material however revealed that Shastasaurus is a short snouted ichthyosaur, and as such much of the long snouted ichthyosaur fossils associated with the type species were found to have actually been placed in the wrong genus, and have since been moved to other genera.
Today Shastasaurus remains are usually associated with the Northern coast lines of the Pacific Ocean, but it needs to be remembered that back in the Triassic today’s continents were still very close to one another and surrounded by a single large ocean.
While the Pacific Rim would have formed much of this coastline, the gradually breaking up land masses created new seas and it is thought by some palaeontologists that Shastasaurus may have ventured into these new areas as well. Shastasaurus also has two dubious species associated with the genus, S. carinthiacus from Austria and S. neubigi from Germany.
Central Europe at the time of the Triassic was more like an Island chain surrounded by shallow seas, not only well away from other Shastasaurus fossil sites, but different from the deep water environments where Shastasaurus is thought to have hunted.
If these remains are not those of Shastasaurus then they may yet represent a similar type of ichthyosaur, whereas if they are the same genus then they may reveal that Shastasaurus was either more flexible in its hunting, or that it was possibly migratory and heading to shallow water birthing areas or moving on to new feeding grounds.
Shastasaurus body features at a glance
Shastasaurus was what is now termed a primitive ichthyosaur. Key to this is what appears to be a lack of a dorsal fin, something which is clearly seen in advanced ichthyosaurs but is lacking in basal forms.
The lack of a dorsal fin bone does not necessarily mean that a dorsal fin was not present; it may have existed in the form of softer tissue such as cartilage which rapidly decomposes in sea water. Still without more direct evidence the wider assumption is that Shastasaurus did not have a dorsal fin.
The tail fluke of Shastasaurus may not have been as well formed as in later ichthyosaurs either. The rear of the body that grew into the tail is seen to have strongly arced downwards indicating that the lower tail lobe was larger and more developed than the upper lobe. This suggests that Shastasaurus was better at cruising rather than active pursuit of prey.
Early art depicting ichthyosaurs often depicts them to have large rounded bodies, and while modern reconstructions suggest that ichthyosaurs in general are much slimmer than what they were classically depicted to be, the ribs in proportion to total body length prove that this was definitely not the case for Shastasaurus.
The distance between fore and aft flippers is roughly seven meters while the ribs were barely two meters tall. This gives Shastasaurus a profile more like that of a torpedo which would have helped to streamline Shastasaurus as it swam through the water.
This effectively means that Shastasaurus faced reduced resistance and experienced less drag so that it did not have to expend so much energy on swimming, something that is especially important given the underdevelopment of the tail fluke.
As mentioned above, the fossil specimens of S. liangae from China allowed for the first accurate reconstruction and analysis of Shastasaurus, particularly the skull. The skull itself is just a little bit over eight per cent of the total length of adult Shastasaurus, and displays extreme shortening adaptations.
While the jaws are reduced, they seem to have reached their minimisation limit as the nasal bone which is usually mid-way in the skull is on the very tip of the jaws. The skull is also laterally compressed which means that when viewed from the front the skull would look narrow like it was pushed in from the sides.
Shastasaurus hunting and feeding specialisation
The addition of new fossil material to create the species S. sikanniensis has made Shastasaurus the largest currently known marine reptile at twenty-one metres long.
This is much larger than others such as the Jurassic predator Liopleurodon which is often incorrectly labelled as the largest and credited as being up to twenty-five meters long, even though the largest fossil specimens for this genus are barely six and half meters long. Shastasaurus is thought to have grown to such a large size so that it was better able to dive to deep depths, as well as being larger than most of the marine predators of the time effectively taking full grown adults off the menu list.
Large size meant that it could take in a greater amount of air and stay down longer than a much smaller ichthyosaur.
Shastasaurus has one of the most specialised mouths of all known ichthyosaurs in that the jaws are both short and toothless. This toothless adaptation is also seen in the similar Shonisaurus although this ichthyosaur has comparatively longer jaws.
Why Shastasaurus should have such short jaws has puzzled many, but analysis of the skulls, particularly the attachments for the jaw opening muscles has given rise to the theory (proposed by M.P. Sander, X. Chen, L. Cheng & X. Wang in 2011) that Shastasaurus was a suction feeder.
Like some fish Shastasaurus opening its mouth wide so fast that it would have created a point where the space and existing water inside its mouth would be reduced to such a low pressure it would create a vacuum inside its mouth.
Water immediately in front of the mouth, as well as anything in it such as squid, would get sucked into this vacuum as the pressure within was equalised. Further evidence for this feeding behaviour comes from the large hyoid bones that would have allowed the tongues to have also been quickly drawn low and back, further increasing the vacuum effect.
The very size of the jaws themselves also support this as shorter jaws would have much less resistance when moving through the water than longer jaws, further increasing the speed that they can be opened which in turn increases the vacuum effect inside the mouth.
If this proposed method of feeding is correct then it’s a wonderful case of nature reverse engineering itself. This is because squid use jets of water pressure squirted out of their bodies to propel themselves through the water, especially when escaping predators. Shastasaurus on the other hand basing its feeding behaviour on using the hydro dynamics of an aquatic vacuum would have denied the squid their advantage, making prey capture for Shastasaurus quite easy.
Further Reading
- – Giant ichthyosaurs of the Triassic – a new species of Shonisaurus from the Pardonet Formation (Norian: Late Triassic) of British Columbia. – Journal of Vertebrate Paleontology 24 (3): 838–849. – E. L. Nicholls & M. Manabe – 2004.
- – On the occurrence of the ichthyosaur Shastasaurus in the Guanling Biota (Late Triassic), Guizhou, China. – Vertebrata PalAsiatica 47 (3): 178–193. – Shang Qing-Hua & Li Chun – 2009.
- – Short-Snouted Toothless Ichthyosaur from China Suggests Late Triassic Diversification of Suction Feeding Ichthyosaurs. – PLoS ONE 6 (5): e19480. – P. M. Sender, X. Chen, L. Cheng & X. Wang – 2011.
- – Absence of Suction Feeding Ichthyosaurs and Its Implications for Triassic Mesopelagic Paleoecology. – PLoS ONE. – R. Motani, T. Tomita, E. Maxwell, D. Jiang & P. Sander – 2013.







