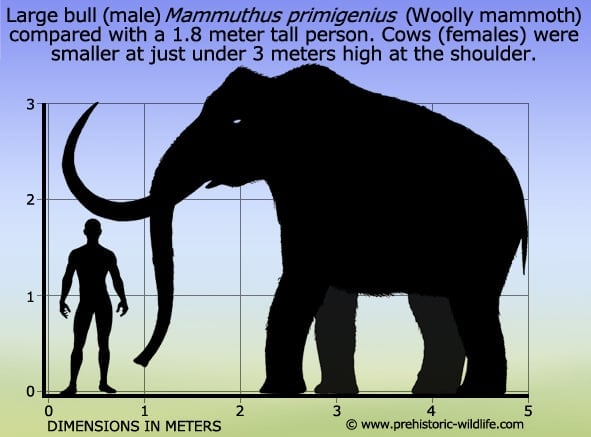
The woolly mammoth of the ice age
Mammuthus primigenius is more popularly known as the woolly mammoth that today is regarded as the poster animal for the ice age, a colloquial term for the Pleistocene period which saw a series of glaciations across the upper latitudes of the Northern hemisphere and an overall reduction in global temperatures.
Sometimes also known as the tundra mammoth, the woolly mammoth is but one species of many mammoths that inhabited northern ecosystems, but the large and often exceptional level of preservation of some remains have revealed more about this prehistoric animal than many others.
The woolly mammoth like all mammoths is closely related to elephants, but features a number of special adaptations that helped it survive in the much colder latitudes of the northern hemisphere.
First and most obvious is the growth of the long shaggy coat of hair over its body, the longest strands of which being up to meter long.
These long hairs covered a denser growth of under hair that provided the main insulation which in turn covered the skin which had a thick layer of fat underneath it to provide even further insulations from the cold.
Still the adaptations went even further as the skin itself had sebaceous glands that would have secreted sebum, an oily substance primarily composed of dead fat cells into the hair. Sebum has a number of functions that help maintain skin and hair integrity, but most important to mammoths is that the secretion of sebum would have helped to waterproof the long hair and further increase its insulatory properties.
Another adaptation to the cold environments is the small size of the ears which are only thirty centimetres long at most, whereas by contrast the African elephant has ears that are up to one hundred and eighty centimetres long. Here it is obvious that the difference is down to thermoregulation.
By having a bigger ears, the African elephant has a larger surface area to lose body heat through, a very useful adaptation for the hot African climate and the reason why African elephants can be frequently observed flapping their large ears. By contrast the woolly mammoth would need to conserve as much body heat as it could, so by having smaller ears it could reduce the surface area to lose heat through.
Woolly mammoths are typically associated with open areas such as grassy plains. The grass on these plains was not especially high in nutritional value but grew in such quantities that it could provide a staple portion of the diet.
Woolly mammoths also seem to have fed upon saplings of trees such as birch which would have helped keep the plains open and covered in grass. Although the cold climate meant that the environment would be dryer, it would still periodically snow and cover the ground.
The tusks of woolly mammoths are thought by many to have grown so long and curved so that they could sweep off the fresh covering of snow and get at the grass underneath, something that has also been proposed for the woolly rhino Coelodonta that was also active in these open habitats at the same time as the woolly mammoth.
The trunk of the woolly mammoth had an upper lip that was larger than the lower, a feature often represented in cave art depicting woolly mammoths that has since been found preserved in at least one frozen specimen. This could have been to help the mammoth to more easily grip hold of tufts of grass while lifting them to the mouth.
The molar teeth of mammoths were large with broad surface areas to provide for efficient processing of large amounts of grass. Like other mammoths M. primigenius had enlarged neural spines rising up from the anterior dorsal vertebrae.
This meant that in life the mammoth would have a hump of soft tissue roughly above where its shoulders were for the purpose of fat storage that allowed the animal to survive during periods where food was not as readily available.
There was probably very little in the way of predators that could take down a fully grown mammoth, but juvenile mammoths seem to have been a regular prey item to big cats similar to how juvenile African elephants are sometimes hunted by African lions today.
In Pleistocene Eurasia however the culprit seems to have been the big cat Homotherium that has been found along with juvenile mammoth remains. Cave Hyenas were also active in these plains environments, but seem to have hunted horses and rhinos instead, although they likely scavenged woolly mammoth carcasses when found.

How woolly mammoths got their colour
In 2006 research conducted by H. Rompler, N. Rohland, C. lalueza-Fox, E. Willerslev, T. Kuznetsova, G. Rabeder, J. Bertranpetit, T. Schoneberg and M. Hofreiter resulted in the extraction of the MC1R (melanocortin 1 receptor) gene from bones.
In simple terms, this is the gene that controls hair colour in the living mammal and like many other known mammals there were dominant and recessive versions of this gene in woolly mammoths. Like in other animals, the dominant gene always overrides the recessive gene when it is present, but when the dominant genes are absent and replaced by other recessive genes, the recessive trait shows through.
The dominant gene was for darker, presumably dark brown hair, while the recessive gene was for lighter hair. Lighter hair could have been to the extent of being blonde or even red/ginger coloured, or simply a pale shade of brown. The image below displays table results of different coloured woolly mammoths that have different sets of genes dependent upon their parents, and is based upon Mendelian genetics. Capitalised ‘M’ is for the dominant gene, lower case ‘m’ is for the recessive.
Both are referred to with the same letter as they affect the same part of the animal, a standard doctrine that prevents possible confusion with other genes although other letters can be used, here this letter represents the MC1R gene. Please remember that two genes in total, one from each parent are passed down to the offspring, the combination resulting in a specific colour, as well as future possibility for further colour mutations.

Table 1 shows an initial paring between a dark and dominant gene only carrier with a light and recessive gene only carrier. The question of how two animals of the same species can have different hair without previous pairing immediately crops up here, but can possibly be best explained by random genetic aberration.
Sometimes animals that are totally dark or totally light (like an albino) are born, and if the progeny survives to reproduce the establishment of differing dominant and recessive genes in a population can quite easily occur. This however is a very basic analogy and is not intended to be an absolute explanation of recessive and dominant genes.
Returning to the subject, the pairing of parent A (both dominant genes) and parent B (both recessive genes) results in all offspring carrying one dominant and one recessive gene. The dominant (dark) gene overrides the recessive (light) so that the mammoth in question has hair colour which is dark, but still carries a copy of the recessive gene.
Table 2 shows a pairing of two mammoths that carry both dominant and recessive genes. Offspring from this pairing are most likely to pick up one of each of the genes to have dark hair but continue to carry the recessive gene themselves.
However due to the genetic mix of the parents there is also a one in four chance the offspring will pick up both dark genes that result in a dark coated mammoth that only carries the dark genes, and a light coated mammoth that only carries recessive genes.
By the law of averages this can also be interpreted as a three in four chance of the offspring having a dark coat, and a one in four chance of it being light.
Tables 3 and 4 show what happens when a mammoth with both genes mates with a mammoth that has only one kind of gene. In table three the result is that all offspring have dark coats, but half have the chance of still carrying the recessive gene. The offspring in table 4 have a fifty-fifty chance of being either light or dark coloured, but all will carry the recessive genes through to the next generation.
Tables 5 and 6 are for scenarios that are termed ‘true breeding’ which means that the outcome of the resulting match is without doubt (barring such things as genetic aberration).
With the resulting principals in mind it is easy to conceive that large populations of woolly mammoths would be mostly dark haired with lighter haired individuals being fewer in number but quite frequently occurring.
More isolated populations however with a reduced gene pool may over the course of successive generations have developed a predominance towards being either mostly dark or light haired, depending upon the original population and survival and reproduction of offspring.
Extinction of Mammuthus primigenius
Woolly mammoths suffered the same fate as most of the Pleistocene megafauna and seem to have become extinct through a combination of climate change and hunting from early humans.
Either one of these two things on their own probably would not have finished the species, but the more factors that change in an animal’s lifestyle, the more said animal has to adapt to the new conditions which decreases its chance of long term survival.
The Pleistocene is marked by a pattern of warmer more temperate time periods that saw ice sheets receding further north, and cooler periods of glaciation where they extended further south.
Woolly mammoths along with their forebears the southern (M. meridionalis) and steppe (M. trogontherii) mammoths coped well with these changes, but the last glaciation towards the end of the Pleistocene seems to have been one of the coldest. Additionally the open areas of grassy plains that mammoths frequented were becoming smaller, being replaced by more densely grown forested areas that would have reduced the amounts of grass that woolly mammoths fed upon.
At the time of these harsh conditions and habitat loss, the relatively new factor of human hunting was also starting to have an effect on mammoth numbers.
With harsher conditions and reduced areas capable of supporting mammoths on the landscape, the overall population of mammoths would decline to a level that the environment could support. This would magnify the impact from hunting from human hunters, as the mammoths no longer had the numbers to cover their losses from hunting.
Eventually the mammoth population would reach a point where they simply could not recover to their previous numbers and would gradually fade away from the landscape.
The disappearance of mammoths was not an overnight event, and actually seems to have occurred over at least several thousand years.
Most of the mammoths seem to have died out on mainland Eurasia at the end of the Pleistocene and early Holocene periods. Small isolated populations did survive on islands for much later however with the most recent currently known being the mammoths of Wrangel Island. A small island in the Arctic Ocean, the tundra here supported woolly mammoths till as recently as 1700BP.

Resurrection through cloning to claims of actual survival
In the story Jurassic Park (the novel and the film adaptation) dinosaurs are brought back to life through retrieval of their DNA. Unfortunately this is pure science fiction as DNA simply does not survive for this long, which means that this method could never be used to resurrect the dinosaurs.
Woolly mammoths however are a very different matter since many of their remains have been found not fossilised like in dinosaurs but actually frozen. This means that the bones and soft tissue have not been mineralised by any fossilisation process but are actually close to what they were in life.
Unfortunately close is not good enough for full extraction of the DNA sequence and at the time of writing the possibility for cloning a mammoth remains a possibility but one that requires significant advancement in the sciences to make real. Part of the problem is the same freezing process that has preserved the remains and the fact that body cells are mostly water.
When water freezes and turns to ice the solid state takes up a larger volume of space (think how when you make ice cubes in the freezer, the cubes always rise up beyond the height of the water when it was liquid).
This means that the water in the cells expands to break through the cellular wall which for lack of a better term destroys the preserved tissue at a cellular level. Additional problems to this are decomposition of remains as well as bacterial and fungal infections that further hamper the preservation of the DNA. However while this makes getting a complete DNA sequence an incredibly difficult task, there are other options available that help scientists to fill the gaps.
Cells are made up of many internal parts, and some in particular are organelles called mitochondria. Mitochondria are usually associated with energy production inside of the cell but they are also store mitochondrial DNA.
Usually abbreviated as mDNA, this is not enough to recreate an animal, but it does allow scientists to identify how closely related an animal is to others, something that could help in identifying a future surrogate animal for an embryo.
Scientists have also tried to extract frozen sperm in the hopes of using it to produce a hybrid with a modern elephant, although even if possible a hybrid would only be half mammoth. The early portion of the twenty-first century has focused upon expanded the methods of viable DNA extraction by using other body parts like hair, with the DNA sequence being built up piece by piece.
The level of success by scientists varies considerably, but new techniques and variations of existing methods have been employed to further the science and increase the chances of success. The wider consensus of the scientific community regarding the resurrection of the woolly mammoth is that it is not so much a question of if, but when it happens.
Even as late as the early twenty-first century there are still claims that small populations of woolly mammoths are still roaming the frozen wastes of Siberia and even Alaska, partly due to the sale of mammoth tusks for the ivory trade.
However despite numerous eye witness reports no living mammoths have been definitely documented, with what little visual evidence in the form of videos and photographs being so fuzzy and blurred identification is impossible. Many pieces of evidence have also in time been proven to be fake, either by in depth analysis of evidence or confessions by the perpetrators of the hoax.
However the regions that would best be able to support woolly mammoths today are some of the most remote and least explored, and it is perhaps not inconceivable that small populations could have survived this far.
Also even if woolly mammoths absolutely do not exist like the majority of scientists believe, these environments may one day see woolly mammoths roaming them again if cloning one day proves so successful that the cloned offspring are released back into the wild.
Further Reading
- – An Account of Elephants Teeth and Bones Found under Ground. – Philosophical Transactions 35 (399–406): 457–471. – H. Sloane – 1727-1728.
- Of Fossile Teeth and Bones of Elephants. Part the Second. – Philosophical Transactions 35 – (399–406): 497–514. – H. Sloane – 1727-1728.
- – A Letter from John Phil. Breyne, M. D. F. R. S. To Sir Hans Sloane, Bart. Pres. R. S. With Observations, and a Description of Some Mammoth’s Bones Dug up in Siberia, Proving Them to Have Belonged to Elephants. – Philosophical Transactions of the Royal Society of London 40 (445–451): 124. – J. P. Breyne & M. Wolochowicz – 1737.
- – M�moire sur les �p�ces d’elephans tant vivantes que fossils, lu � la s�ance publique de l’Institut National le 15 germinal, an IV. – Magasin encyclop�dique, 2e an�e 440–445. G. Cuvier – 1796.
- – A Contribution to the Morphology of the Mammoth, Elephas Primigenius Blumenbach; With an Explanation of My Attempt at a Restoration. – Annual report of the Board of Regents of the Smithsonian Institution: 326–334. – E. Pfitzenmayer – 1907.
- – Frozen Mammoths and Modern Geology: The death of the giants can be explained as a hazard of tundra life, without evoking catastrophic events. – Science 133 (3455): 729–735. – W. R. Farrand – 1961.
- – Hair structure of the Woolly mammoth, Mammuthus primigenius and the modern elephants, Elephas maximus and Loxodonta africana. – Journal of Zoology 199 (2): 271–274. A. Valente – 1983.
- – Holocene dwarf mammoths from Wrangel Island in the Siberian Arctic. – Nature 362 (6418): 337. – S. L. Vartanyan, V. E. Garutt & A. V. Sher – 1993.
- – Contribution of isotopic biogeochemistry (13C,15N,18O) to the paleoecology of mammoths (Mammuthus primigenius). – Historical Biology 7 (3): 187. – H. Bocherens, M. Fizet, A. Mariotti, R. A. Gangloff, J. A. Burns – 1994.
- – The latest woolly mammoths (Mammuthus primigenius Blumenbach) in Europe and Asia: A review of the current evidence. – Quaternary Science Reviews 21 (14–15): 1559 – A. J. Stuart, L. D. Sulerzhitsky, L. A. Orlova, Y. V. Kuzmin, A. M. Lister – 2002.
- – Structure and evolution of mammoth molar enamel. – Acta Palaeontologica Polonica. 3 48: 383–396. M. P. Ferretti – 2003.
- – Radiocarbon evidence of mid-Holocene mammoths stranded on an Alaskan Bering Sea island. – Nature 429 (6993): 746–749. – r. Dale Guthrie – 2004.
- – Pleistocene to Holocene extinction dynamics in giant deer and woolly mammoth. – Nature 431 (7009): 684–689.- A. J. Stuart, P. A. Kosintsev, T. F. G. Higham, A. M. Lister – 2004.
- – Sebaceous Glands of the Woolly Mammoth, Mammothus primigenius Blum.: Histological Evidence. – Doklady Biological Sciences 398: 382–384. – V. E. Repin, O. S. Taranov, E. I. Ryabchikova, A. N. Tikhonov, V. G. Pugachev – 2004.
- – Advances in proboscidean taxonomy & classification, anatomy & physiology, and ecology & behavior. – Quaternary International. 126–128 – J. Shoshani & P. Tassy – 2005.
- – 5,700-Year-Old Mammoth Remains from the Pribilof Islands, Alaska: Last Outpost of North America Megafauna. – Geological Society of America 37: 463. – K. S. Crossen – 2005.
- – Multiplex amplification of the mammoth mitochondrial genome and the evolution of Elephantidae. Nature 439 (7077): 724–727. – J. Krause, P. H. Dear, J. L. Pollack, M. Slatkin, H. Spriggs, I. Barnes, A. M. Lister, Ebersberger, S. P��bo, M. Hofreite – 2005.
- – The pattern and process of mammoth evolution in Eurasia. – Quaternary International. 126–128: 49. – A. M. Lister, A. V. Sher, H. Van Essen, G. Wei – 2005.
- – Mammoth tracks indicate a declining Late Pleistocene population in southwestern Alberta, Canada. – Quaternary Science Reviews 24 (10–11): 1253. – P. McNeil, L. Hills, B. Kooyman, S. Tolman – 2005.
- – The extinction of woolly mammoth (Mammuthus primigenius) and straight-tusked elephant (Palaeoloxodon antiquus) in Europe. – Quaternary International. 126–128: 171–177. – A. J. Stuart – 2005.
- – Nuclear Gene Indicates Coat-Color Polymorphism in Mammoths. – Science 313 (5783): 62. – H. Rompler, N. Rohland, C. Lalueza-Fox, E. Willerslev, T. Kuznetsova, G. Rabeder, J. Bertranpetit, T. Sch�neberg, M. Hofreiter – 2006.
- – Reading the Evolutionary History of the Woolly Mammoth in Its Mitochondrial Genome. – PLoS Biology 4 (3): e74. L. Gross – 2006.
- – AMS 14C chronology of the world’s southernmost woolly mammoth (Mammuthus primigenius Blum.). – Quaternary Science Reviews 26 (7–8): 954. – K. Takahashi, G. Wei, H. Uno, M. Yoneda, C. Jin, C. Sun, S. Zhang, B. Zhong – 2007.
- – Intraspecific phylogenetic analysis of Siberian woolly mammoths using complete mitochondrial genomes. – Proceedings of the National Academy of Sciences 105 (24): 8327. – M. T. P. Gilbert, D. I. Drautz, A. M. Lesk, S. Y. W. Ho, J. Qi, A. Ratan, C-H. Hsu, A. Sher, L. Dalen, A. Gotherstrom, L. P. Tomsho, S. Rendulic, M. Packard, P. F. Campos, T. V. Kuznetsova, F. Shidlovskiy, A. Tikhonov, E. Willerslev, P. Iacumin, B. Buigues, P. G. P. Ericson, M. Germonpre, P. Kosintsev, V. Nikolaev, M. Nowak-Kemp, J. R. Knight, G. P. Irzyk, C. S. Perbost, K. M. Fredrikson, T. T. Harkins – 2008.
- – Out of America: Ancient DNA Evidence for a New World Origin of Late Quaternary Woolly Mammoths. – Current Biology 18 (17): 1320–1326. – R. Debruyne, G. Chu, C. E. King, K. Bos, M. Kuch, C. Schwarz, P. Szpak, D. R. Gr�cke, P. Matheus, G. Zazula, D. Guthrie, D. Froese, B. Buigues, C. De Marliave, C. Flemming, D. Poinar, D. Fisher, J. Southon, A. N. Tikhonov, R. D. E. MacPhee, H. N. Poinar – 2008.
- – Collection of radiocarbon dates on the mammoths (Mammuthus primigenius) and other genera of Wrangel Island, northeast Siberia, Russia. – Quaternary Research 70: 51. – S. L. Vartanyan, K. A. Arslanov, J. A. Karhu, G. R. Possnert, L. D. Sulerzhitsky – 2008.
- – Single carcass of Mammuthus primigenius with lithic artifacts in the Upper Pleistocene of northern Italy. – Journal of Archaeological Science 35 (9): 2606 – M. Mussi & P. Villa – 2008.
- – Climate Change, Humans, and the Extinction of the Woolly Mammoth. – In Barnosky, Anthony. PLoS Biology 6 (4): e79. – D. Nogu�s-Bravo, J. S. Rodr�guez, J. N. Hortal, P. Batra & M. B. Ara�jo – 2008.
- – What Killed the Woolly Mammoth? – PLoS Biology 6 (4): e99 – C. Sedwick – 2008.
- – Temporal genetic change in the last remaining population of woolly mammoth. – Proceedings of the Royal Society B: Biological Sciences 277 (1692): 2331. – V. Nystrom, L. Dalen, S. Vartanyan, K. Liden, N. Ryman, A. Angerbjorn – 2010.
- – Genomic DNA Sequences from Mastodon and Woolly Mammoth Reveal Deep Speciation of Forest and Savanna Elephants. – In Penny, David. PLoS Biology 8 (12): e1000564. – N. Rohland, D. Reich, S. Mallick, M. Meyer, R. E. Green, N. J. Georgiadis, A. Roca, M. – Hofreiter – 2010.
- – Substitutions in woolly mammoth hemoglobin confer biochemical properties adaptive for cold tolerance. – Nature Genetics 42 (6): 536–540. – K. L. Campbell, J. E. E. Roberts, L. N. Watson, J. R. Stetefeld, A. M. Sloan, A. V. Signore, J. W. Howatt, J. R. H. Tame, N. Rohland, T. J. Shen, J. J. Austin, M. Hofreiter, C. Ho, R. E. Weber & A. Cooper – 2010.
- – Nursing, weaning, and tooth development in woolly mammoths from Old Crow, Yukon, Canada: Implications for Pleistocene extinctions. – Palaeogeography, Palaeoclimatology, Palaeoecology 298 (3–4): 257. J. Z. Metcalfe, F. J. Longstaffe, G. D. Zazula – 2010.
- – Palaeo-environmental and dietary analysis of intestinal contents of a mammoth calf (Yamal Peninsula, northwest Siberia). – Quaternary Science Reviews 30 (27–28) – B. Van Geel, D. C. Fisher, A. N. Rountrey, J. Van Arkel, J. F. Duivenvoorden, A. M. Nieman, G. B. A. Van Reenen, A. N. Tikhonov, B. Buigues, B. Gravendeel – 2011.
- – Complete Columbian mammoth mitogenome suggests interbreeding with woolly mammoths. – Genome Biology 12 (5): R51. – J. Enk, A. Devault, R. Debruyne, C. E. King, T. Treangen, D. O’Rourke, S. L. Salzberg, D. Fisher, R. MacPhee, H. Poinar – 2011.
- – Population-level genotyping of coat colour polymorphism in woolly mammoth (Mammuthus primigenius). – Quaternary Science Reviews 30 (17–18): 2304. – C. Workman, L. Dalen, S. Vartanyan, B. Shapiro, P. Kosintsev, A. Sher, A. Gotherstrom, I. Barnes – 2011.
- – Anatomy, death, and preservation of a woolly mammoth (Mammuthus primigenius) calf, Yamal Peninsula, northwest Siberia. – Quaternary International 255: 94. – D. C. Fisher, A. N. Tikhonov, P. A. Kosintsev, A. N. Rountrey, B. Buigues, J. Van Der Plicht – 2012.
- – Mammuthus primigenius in the cave and portable art: An overview with a short account on the elephant fossil record in Southern Europe during the last glacial. Quaternary International. 276-277: 61. – I. M. Braun & M. R. Palombo – 2012.
- – Environmental reconstruction inferred from the intestinal contents of the Yamal baby mammoth Lyuba (Mammuthus primigenius Blumenbach, 1799). – Quaternary International 255: 231. – P. A. Kosintsev, E. G. Lapteva, S. S. Trofimova, O. G. Zanina, A. N. Tikhonov & J. Van Der Plicht – 2012.
- – Early tooth development, gestation, and season of birth in mammoths. – Quaternary International 255: 196. – A. N. Rountrey, D. C. Fisher, A. N. Tikhonov, P. A. Kosintsev, P. A. Lazarev, G. Boeskorov, B. Buigues – 2012.
- – Proteomic analysis of a Pleistocene mammoth femur reveals more than one hundred ancient bone proteins. – Journal of Proteome Research 11 (2): 917–926. – E. Cappellini, L. J. Jensen, D. Szklarczyk, A. L.Ginolhac, R. A. R. Da Fonseca, T. W. Stafford, S. R. Holen, M. J. Collins, L. Orlando, E. Willerslev, M. T. P. Gilbert, J. V. Olsen – 2012.
- – Microsatellite genotyping reveals end-Pleistocene decline in mammoth autosomal genetic variation. – Molecular Ecology 21 (14): 3391–3402. – V. Nystr�m, J. Humphrey, P. Skoglund, N. J. McKeown, S. Vartanyan, P. W. Shaw, K. Lid�n, M. Jakobsson, I. A. N. Barnes, A. Angerbj�rn, A. Lister, L. Dal�n – 2012.
- – Did mammoth end up a lilliput? Temporal body size trends in Late Pleistocene Mammoths, Mammuthus primigenius (Blumenbach, 1799) inferred from dental data. – Quaternary International 255: 53. – N. Den Ouden, J. W. F. Reumer, L. W. Van Den Hoek Ostende – 2012.
- – Comparative revision of the Iberian woolly mammoth (Mammuthus primigenius) record into a European context. – Quaternary Science Reviews 32: 64. – D. J. �lvarez-Lao & N. Garc�a – 2012.
- – Can a mammoth carcass really preserve flowing blood and possibly live cells? – Nature. – K. Wong – 2013.
- – X-ray computed tomography of two mammoth calf mummies. – Journal of Paleontology. 88 (4): 664–675. – Daniel C. Fisher – 2014.
- – Megafaunal split ends: microscopical characterisation of hair structure and function in extinct woolly mammoth and woolly rhino. – Quaternary Science Reviews. 83: 68–75. – Silvana R. Tridico, Paul Rigby, Paul Kirkbride, James Haile & Michael Bunce – 2014.
- – Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths. – PeerJ. 2: e318. – J. W. F. Reumer, C. M. A. Ten Broek & F. Galis – 2014.
- – New data on trunk morphology in the woolly mammoth, Mammuthus primigenius (Blumenbach). – Paleontological Journal. 49 (2): 200–210. – V. V. Plotnikov, E. N. Maschenko, I. S. Pavlov, A. V. Protopopov, G. G. Boeskorov & E. A. Petrova – 2015.
- – Complete Genomes Reveal Signatures of Demographic and Genetic Declines in the Woolly Mammoth. – Current Biology. 25 (10): 1395–1400. – Eleftheria Palkopoulou, Swapan Mallick, Pontus Skoglund, Jacob Enk, Nadin Rohland, Heng Li, Ay�a Omrak, Sergey Vartanyan, Hendrik Poinar, Anders G�therstr�m, David Reich & Love Dal�n1 – 2015.
- – Elephantid genomes reveal the molecular bases of Woolly Mammoth adaptations to the arctic. – Cell Reports. 12 (2): 217–228. – V. Lynch, O. C. Bedoya-Reina, A. Ratan, M. Sulak, D. I. Drautz-Moses, G. H. Perry, W. Miller & S. C. Schuster – 2015.
- – Signals of positive selection in mitochondrial protein‐coding genes of woolly mammoth: Adaptation to extreme environments?. – Ecology and Evolution. 9 (12): 6821–6832. doi:10.1002/ece3.5250. – Jacob Njaramba Ngatia, Tian Ming Lan, Thi Dao Dinh, Le Zhang, Ahmed Khalid Ahmed & Yan Chun Xu – 2019.
- – Mammoth ivory was the most suitable osseous raw material for the production of Late Pleistocene big game projectile points. – Scientific Reports. 9 (1): 2303. – S. J. Pfeifer, W. L. Hartramph, R.-D. Kahlke & F. A. M�ller – 2019.
- – Thriving or surviving? The isotopic record of the Wrangel Island woolly mammoth population. – Quaternary Science Reviews. 222: 105884. – L. Arppe, J. A. karhu, S. Vartanyan, D. G. Drucker, H. Etu-Sihvola & H. Bocherens – 2019.
- – Signs of biological activities of 28,000-year-old mammoth nuclei in mouse oocytes visualized by live-cell imaging. – Scientific Reports. 9 (1): 4050. – Kazuo Yamagata, Kouhei Nagai, Hiroshi Miyamoto, Masayuki Anzai, Hiromi Kato, Kei Miyamoto, Satoshi Kurosaka, Rika Azuma, Igor I. Kolodeznikov, Albert V. Protopopov, Valerii V. Plotnikov, Hisato Kobayashi, Ryouka Kawahara-Miki, Tomohiro Kono, Masao Uchida, Yasuyuki Shibata, Tetsuya Handa, Hiroshi Kimura, Yoshihiko Hosoi, Tasuku Mitani, Kazuya Matsumoto & Akira Iritani – 2019.
- – Big tuskers: Maximum sizes of tusks in woolly mammoths – Mammuthus primigenius (Blumenbach) – from East Siberia. – Quaternary International. 537: 88–96. – G. Boeskorov, A. Tikhonov, M. Shchelchkova, J. P. Ballard & D. Mol – 2020.








