Lambeosaurus the dinosaur
Lambeosaurus is one of the hadrosaurs that is frequently depicted in dinosaur books and toys, and study of this genus is required learning for anyone who wishes to know more about the hadrosaurs in general.
Lambeosaurus is best known from Canada and upper Montana USA, although other attributed fossils in the past have suggested that Lambeosaurus had a significantly broader range.
The most distinctive feature of Lambeosaurus is the crest which is often described as being either hatchet or axe shaped, the broader forward portion being the blade while the thin rearward projection is the handle.
This rear projection is solid; however the forward portion is hollow with a network of passages inside (a signature feature of the lambeosaurine hadrosaurids).
The function of the crest is one of the most often talked about areas of the body.
One explanation is that the crest was used as either a snorkel or an air chamber for trapping a supply of breathable air so that Lambeosaurus could hold their heads underwater for extended periods.
This however would require Lambeosaurus to regularly be active within bodies of water, and it does not explain the variance in form of head crests between Lambeosaurus and other lambeosaurine genera.
One exciting theory is that the crests allowed for the storage of salt glands, special organs that allow excess salt from the environments to be excreted.
However the presence of salt glands is usually associated with animals that live in marine (saltwater) ecosystems which have higher levels of salts, thus necessitating the presence of salt glands.
This is not actually that much of a stretch to believe however since during the late Cretaceous most of central North America was submerged by a shallow sea called the Western Interior Seaway that ran North to South from what is now Upper Canada into the Gulf of Mexico.
The areas that Lambeosaurus remains are known from have been interpreted as being marshes and swamps that were in close proximity to the coastline of the Western Interior Sea. Additionally a 1979 paper by palaeontologist Jack R. Horner described what appeared to be Lambeosaurus jaws (L. magnicristatus) from marine sediment.
The problems with this theory however is that it is that with this evidence it is only relevant to Lambeosaurus, with many other hadrosaurs being known from different ecosystems.
Additionally this does not explain the difference in crest form between not only hadrosaur genera, but the two main Lambeosaurus species.
The only theory that explains the different crest forms is that of display.
Firstly different species of Lambeosaurus would be able to tell each other apart by the different form of head crest, as well as any hadrosaurid other genera that were active in the same locations as they were.
Additionally the crests of adult Lambeosaurus were more developed than juveniles which indicates that they were also a sign of maturity.
Specimens thought to be females also have more rounded crests than the males.
Another factor concerning display is that of sound.
The forward portion of the crest is hollow with a network of tubes, and assuming this is not just a weight saving feature to reduce stress on the head and neck, this may have formed a resonating chamber for amplifying the calls.
Not only have studies continually revisited this area, but different crest variations between species should allow for different sounding calls.
Study in one specimen of the related Corythosaurus also suggests that lambeosaurine hadrosaurids in particular had very good hearing, further supporting the auditory hypothesis for the crest.
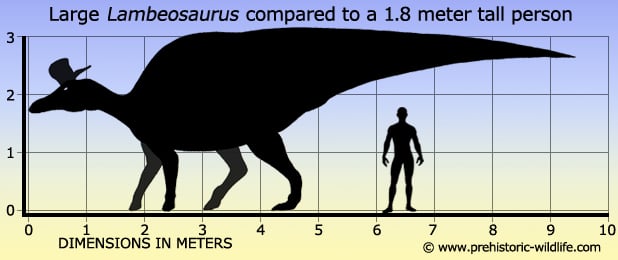
Size estimates for Lambeosaurus can vary greatly depending upon source because of the large amounts of fossil remains that have been attributed to the genus.
The confirmed Lambeosaurus remains that are known from Alberta Canada and Montana, USA point to individuals around the nine to nine and a half meter mark for total length.
Upper estimates of fifteen meters long still exist in some sources however, though this estimate is based upon material that is no longer attributed to the Lambeosaurus genus.
This stems back to the description of a dubious species of Lambeosaurus in 1981 called L. laticaudus from fossils found in Baja California.
The Lambeosaurus connection however could never be confirmed since the crest is missing.
In 2012 these remains were re-established as a new genus called Magnapaulia.
As with all hadrosaurids, Lambeosaurus seems to have been primarily quadrupedal with the ability to balance and possibly walk on just the rear legs when it had to.
Out of the original five digits of the fore-arm hand, the first digit is missing, lost through selective evolutionary factors.
Digits two, three and four are developed into weight bearing hooves, and when combined with footprints of other hadrosaurids, confirms the idea that Lambeosaurus, and other hadrosaurids, were mostly bipedal.
The fifth digit is free from the others and may have been to help grip and/or balance when the body weight was shifted forward.
By being both quadrupedal and occasionally bipedal, Lambeosaurus could browse upon anything from low to moderately high growing vegetation.
The anterior portion of the mouth however is narrower than in many other hadrosaurids, something that suggests that Lambeosaurus were more selective in their browsing habits.
Once food was removed from the main plant, it was then processed by the grinding teeth that were arranged in batteries at the back of the mouth.
Lambeosaurus possibly shared its habitat with other genera of hadrosaur such as Corythosaurus, though this genus is currently known from slightly older Campanian deposits, which indicates that Lambeosaurus may have actually replaced Corythosaurus.
Other dinosaurs roughly active around that time and locale include the ceratopsians Chasmosaurus and Centrosaurus, the nodosaur Edmontonia as well as the ankylosaur Euoplocephalus.
However the exact relationships between these different dinosaurs is still being established since some such as ankylosaurs seem to have been better adapted to dryer more arid environments due to the arrangements of their nasal passages.
Predators of Lambeosaurus may have included Campanian era tyrannosaurs such as Albertosaurus and Daspletosaurus.
Classification history of Lambeosaurus
The history of Lambeosaurus is one that is steeped in the misinterpretation of dinosaur fossils at a time when palaeontology as we know it today was still in its infancy.
The first Lambeosaurus remains were described by Lawrence Lambe in 1902, however these fossils were only of partial post cranial remains, and rather than establish them as a new genus, Lambe assigned them as a species of Trachodon, T. marginatus.
The problem here is that Trachodon has always been a very dubious genus.
Back in 1856 another palaeontologist named Joseph Leidy created the Trachodon dinosaur genus based only upon the description of a collection of teeth.
Today we now know this description to be flawed since the teeth assigned to Trachodon are actually a combination of both hadrosaur and ceratopsian teeth, two distantly related yet very different groups of dinosaurs.
Leidy named many genera of prehistoric animals many from only teeth and some like Aublysodon remain very dubious, while others have been cast down as valid genera.
But Leidy did achieve one very big success from only describing teeth with his naming of Troodon in 1856.
The dubious nature of Trachodon seems to have still been appreciated by Lambe since when he described two new skulls in 1914 that he thought belonged to T. marginatus he created the genus Stephanosaurus.
The problem here is the same as many extinct animals that have their initial descriptions based upon very incomplete remains.
The fossils originally described as T. marginatus were of the post cranial skeleton with no elements of the skull.
The original concept of the skulls being placed with this material is based around the fact that the skulls came from roughly the same area as the T. marginatus fossils.
This makes the assumptions that they belong to the same genus possible, but one that is impossible to establish without a set of remains that bridge the gaps between both groups of fossils (i.e., skull and post cranial fossils of one individual that can be compared to existing fossils assigned to the genus).
This is why in 1923 William parks removed the skulls from Stephanosaurus (today regarded as a dubious genus) and redescribed them as a new genus, Lambeosaurus, in honour of Lawrence Lambe who had died four years earlier in 1919.
For many years Lambeosaurus (including its earlier association with Stephanosaurus) was not known to have very many fossils.
Today however we now know that that was never the case, further Lambeosaurus were actually being discovered in the early years of the twentieth century.
In 1917 a new dinosaur called Cheneosaurus was named by Lawrence Lambe, a dinosaur that would become the type genus of the Cheneosaurinae, a group of dinosaurs that resembled hadrosaurs, but were much smaller.
Before this in 1902, Henry Fairfield Osborn named a genus Didanodon altidens based upon a partial left jaw.
Lambe later re-classed this specimen as a species of Trachodon, T. altidens.
In 1920 William Diller Matthew took a photograph of the skeletal remains of one such dinosaur and named it Procheneosaurus; however this is not the usual method for establishing a new genus.
When William parks described the genus Tetragonosaurus from other fossils recovered from the Dinosaur Park Formation, he also included the fossils of Procheneosaurus into the type species of Tetragonosaurus, T. praeceps (A second species of T. erectofrons was established for smaller specimens).
The Tetragonosaurus genus would grow further with the addition of T. cranibrevis by Charles M. Sternberg in 1935.
However, nearly forty years’ worth of classification was turned on its head in 1942 when Richard Swann Lull and Nelda Wright released a detailed study about these and associated dinosaurs.
In this study they declared that the whole Tetragonosaurus genus should actually be called Procheneosaurus, as well as possibly Trachodon altidens too.
The really ground breaking discovery however would come in 1975 when Peter Dodson demonstrated that the cheneosaurs were actually made up of the juveniles of the larger hadrosaurs.
Specifically the type specimen of what was now Procheneosaurus was seen to now be a juvenile Lambeosaurus, while other specimens were probably juveniles of Corythosaurus.
The problem here is that technically Procheneosaurus was named before Lambeosaurus, and usually the older name takes precedence over any future naming.
Despite this Lambeosaurus is still credited as being the specific genus for these remains with Procheneosaurus being treated as a synonym.
Another case of a dinosaur retaining a second name despite having fossils attributed to an earlier description of another genus is that of Tyrannosaurus.
Today Lambeosaurus is best known for being the type genus of the Lambeosaurinae, one of the two largest groups of hadrosaurs.
Members of this group, referred to as lambeosaurines, all exhibit hollow head crests, though the physical form of the crest can vary greatly between different genera.
Other lambeosaurine genera closely related to Lambeosaurus include Olorotitan, Corythosaurus and Hypacrosaurus.
Corythosaurus in particular is very similar in physical form to Lambeosaurus, so much so that the only obvious differences between them is the shape and form of the crest and how it attaches to the skull.
There were once seven species of Lambeosaurus, but at the time of writing only the type species L. lambei and L. magnicristatus are universally considered to be valid.
Both of these species are known from upper North America and can be distinguished on the basis of different crest form.
L. lambei has a rear bony projection that is roughly the same length as the forward crest, while L. magnicristatus has a reduced bony projection and a slightly enlarged forward crest.
Another species called L. paucidens is only known from post cranial remains so it is hard to be certain how or even if it is related to the other two Lambeosaurus species.
With this in mind, many palaeontologists consider these remains to instead be a species of Hadrosaurus (H. paucidens).
Other past species have since been established to be different sexes of existing hadrosaur genera, while others (such as L. laticaudus stated above) are now different genera.
Further Reading
- - Corythosaurus intermedius, a new species of trachodont dinosaur, William A. Parks - 1923.
- - A new genus and two new species of trachodont dinosaurs from the Belly River Formation of Alberta, William A. Parks - 1931.
- - New species of trachodont dinosaurs from the Cretaceous formations of Alberta, William A. Parks - 1931.
- - Taxonomic implications of relative growth in lambeosaurine dinosaurs, Peter Dodson - 1975.
- - Upper Cretaceous dinosaurs from the Bearpaw Shale (marine) of south-central Montana with a checklist of Upper Cretaceous dinosaur remains from marine sediments in North America, Jack R. Horner - 1979.
- - A new species of hadrosaurian dinosaur from the Upper Cretaceous of Baja California: ?Lambeosaurus laticaudus, William J. Morris - 1981.
- - Acoustic analyses of potential vocalization in lambeosaurine dinosaurs, David B. Weishampel - 1981.
- - Anatomy and relationships of Lambeosaurus magnicristatus, a crested hadrosaurid dinosaur (Ornithischia) from the Dinosaur Park Formation, Alberta, David C. Evans & Robert R. Reisz - 2007.








