In Depth
Thescelosaurus seems to have been one of the main herbivorous dinosaurs roaming around North America towards the end of the Cretaceous period. The first fossils of Thescelosaurus began to be recovered in the closing decade of the nineteenth century, but were not described until 1913 by Charles W. Gilmore, who provided a much more detailed description in 1915. For over a hundred years afterwards numerous individuals of Thescelosaurus were recovered from across the central portion of North America, resulting in three distinct species being named. One former genus called Bugenasaura has also been discovered to be a junior synonym to Thescelosaurus. Another genus named Parksosaurus that in the past has been speculated to be synonymous with Thescelosaurus, has in recent times been re-affirmed as a distinct genus.
Thescelosaurus has often been described as a hypsilophodont, but modern analysis of this dinosaur has found that Thescelosaurus is actually quite different from Hypsilophodon, and therefore not likely related beyond both of them being ornithopod dinosaurs. Like with most other ornithopod dinosaurs, Thescelosaurus was a bipedal dinosaur which had a body counterbalanced by a long tail that accounted for much of the total body length. Bizarrely for an ornithopod dinosaur, the upper leg of Thescelosaurus was longer that the lower leg, the complete opposite to the arrangement seen in most other ornithopods. This meant that Thescelosaurus would have actually been a relatively slow runner. This raises questions as to how Thescelosaurus survived since there are no other obvious defensive features on this dinosaur, yet we know that Thescelosaurus became relatively widespread in North America. Scutes found in association with one Thescelosaurus specimen may indicate that at least one line of bony armour was present on the body, however an alternative theory is that these scutes may have come from a crocodile since these scutes have so far not been found with any other individuals.
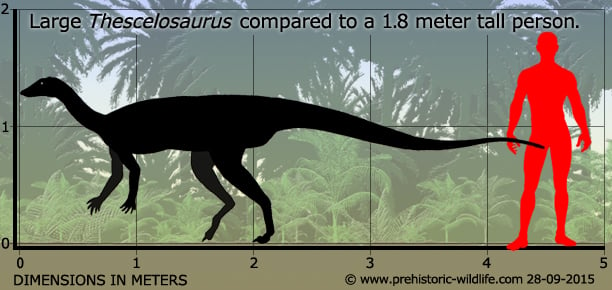
Numerous individuals of Thescelosaurus are now known to us, but there seems to have been a wide variance in the sizes of adults which usually range between two and half and four meters in length. Some of this difference is likely down to a species level, with individuals of T. garbanii often approaching four meters in length, though one specimen seems to have been at least four and a half meters long. It’s possible though that Thescelosaurus may have been sexually dimorphic, meaning that there was a clear difference in how large males and females grew.
Thescelosaurus had pointed teeth at the front of the mouth and leaf shaped teeth towards the back. Leaf shaped teeth like these are common in herbivorous dinosaurs that feed upon softer plants as they easily slice through leafy plant material. The pointed teeth are a little more puzzling however, as they are not as well suited to an exclusive diet of plants. This has led some to speculate that Thescelosaurus might have been omnivorous, either eating small animals like lizards or occasionally scavenging carrion. Prominent ridges on the maxilla bones of the skull and the observation that the leaf-shaped maxilla teeth are set well inside the mouth support the idea that Thescelosaurus had quite muscular cheeks to stop food spilling out of the sides of the mouth when processing food. Six pairs of small teeth were also present in the pre-maxilla, though the tip of the premaxilla was toothless to accommodate the horny beak that covered the front of the mouth. This beak would have been the primary shearing apparatus when cropping plants.
There are two schools of thought concerning which kind of habitat Thescelosaurus preferred. One is that Thescelosaurus preferred channel systems, feeding upon the banks of rivers and streams, while another is that Thescelosaurus preferred floodplains. It may not sound like there is a lot of difference, and indeed the two habitats are connected. Both have an abundance of plants that grow in fertile soil, but channel systems feature a more regular growth of plants where growth on floodplains can be more seasonal. Most Thescelosaurus fossils have been dug out of sandstone, which is more a sign of a channel environment, whereas floodplains tend to produce mudstone. There are also no mass bone beds of Thescelosaurus individuals, again indicating a lack of flooding victims.
If Thescelosaurus lived near water browsing upon softer plants growing by the water, then this may have also been an escape route for them when they got attacked. As soon as a predator showed up they may have entered and swum into and across the river, explaining why Thescelosaurus did not retain leg proportions for running, and also how the genus came to be so widespread. Reduced limb proportions would have also helped with locomotion through the water by not being so easy to get tangled in underwater weeds and debris, and if the latest reconstructions of Spinosaurus are anything to go by, then limb reduction in semi-aquatic dinosaurs may have been an easy thing to achieve.
Thescelosaurus lived during the latest stage of the Cretaceous and lived in the same locations as hadrosaurs, ceratopsians, ankylosaurs, pachycephalosaurs and ornithomimids. Predatory threats to Thescelosaurus were many and included the famous tyrannosaurs like Tyrannosaurus, to troodonts like Troodon, to one of the largest dromaeosaurs, Dakotaraptor. There has been a lot of debate concerning a possible ‘heart’ believed to have been fossilised in an individual Thescelosaurus that was found in 1993. In a paper by Fisher et al published in 2000, they described a fossilised structure found within a specimen of Thescelosaurus that resembled a heart. Their theory was that the original heart muscle had been saponified (turned to soap) as a result of anaerobic conditions and then through regular fossilisation turned to geothite. The authors of this paper noted that the ‘heart’ was comparable with the heart of an ostrich with four chambers, and that Thescelosaurus and probably other dinosaurs had an elevated metabolic rate closer to what we would term as warm-blooded.
It was not long after that doubts about the interpretation of this ‘heart’ began to be publically voiced. The main problem that many people had was that the ‘heart’ just did not look right with anomalies in the basic structure. In a 2001 paper by Timothy et al the ‘heart’ was re-interpreted as a simple concretion that had simply built up an extra dense portion of sedimentary rock. It was also noted that the ‘heart’ partially engulfed one of the ribs and that a second similar concretion could be found behind the right leg. Also observed was the part preserved as the aorta, the main blood vessel that should carry oxygenated blood from the heart. Many noted that this aorta actually became narrower where it actually joined the ‘heart’, something unprecedented in heart anatomy as this would restrict blood flow. There were also no arteries branching off from this aorta to take blood to other parts of the body. Despite these observations, the original authors who described the ‘heart’ defended their earlier work, saying that the ‘heart’ was a concretion, but one built up to the structure of the original heart and aorta.
In 2011 a new study of the ‘heart’ by Cleland, Stoskopf and Schweitzer used state of the art investigative methods including X-ray diffraction, X-ray photoelectron spectroscopy, scanning electron microscopy, histology and CT scanning to determine the truth of the matter. Actually using computer modelling to ‘see inside’ the ‘heart’, they revealed that this structure did not have four chambers like it was previously expected to. Instead they mapped out three areas filled with less dense material than the surrounding ‘heart’ structure. Furthermore, these areas were completely unconnected, which means in absolutely no way could they be heart chambers, as heart chambers need to be connected so that blood can flow through the heart.
The 2011 study also failed to find any of the common elements that are found in fossilised remains of biological structures (i.e. carbon, nitrogen, phosphorus). This means that this ‘heart’ was composed of inorganic material that was not part of the original dinosaur. There was however one patch where the impressions of soft tissues identifiable at the cellular level were observed. These impressions though do not reflect a heart structure, and were recorded as simply being a case of a concretions pressing against soft tissues and then preserving the form while hardening.
The overall conclusion is that this structure is simply a concretion of sand hardened to stone in such a way that at a glance it has become a simulacrum of a heart, not the actual fossilised remains of one.
Further Reading
- A new dinosaur from the Lance Formation of Wyoming. -Smithsonian Miscellaneous Publications 61(5):1-5. - Charles W. Gilmore - 1913. - Osteology of Thescelosaurus, an orthopodus dinosaur from the Lance Formation of Wyoming. - Proceedings of the U.S. National Museum 49 (2127): 591–616. - Charles W. Gilmore - 1915. - Thescelosaurus warreni, a new species of orthopodous dinosaur from the Edmonton Formation of Alberta. - University of Toronto Studies (Geological Series) 21: 1–42. - William A. Parks - 1926. - Classification of Thescelosaurus, with a description of a new species. - Geological Society of America Proceedings for 1936: 365. - Charles M. Sternberg - 1937. - Thescelosaurus edmontonensis, n. sp., and classification of the Hypsilophodontidae. - Journal of Paleontology 14 (5): 481–494. - Charles M. Sternberg - 1940. - Notes on Thescelosaurus, a conservative ornithopod dinosaur from the Upper Cretaceous of North America, with comments on ornithopod classification. - Journal of Paleontology 48 (5): 1048–1067. - Peter M. Galton - 1974. - Dinosaurs of the Lance Formation in eastern Wyoming, by Kraig Derstler - In, The Dinosaurs of Wyoming. Wyoming Geological Association Guidebook, 44th Annual Field Conference. Wyoming Geological Association. pp. 127–146. - Gerald E. Nelson (ed.) 1994. - The species of the basal hypsilophodontid dinosaur Thescelosaurus Gilmore (Ornithischia: Ornithopoda) from the Late Cretaceous of North America. - Neues Jahrbuch f�ur Geologie und Pal�ontologie Abhandlungen 198 (3): 297–311. - Peter M. Galton - 1995. - Cranial anatomy of the basal hypsilophodontid dinosaur Thescelosaurus neglectus Gilmore (Ornithischia; Ornithopoda) from the Upper Cretaceous of North America. - Revue Pal�obiologie, Gen�ve 16 (1): 231–258. - Peter M. Galton - 1997. - Cranial anatomy of the hypsilophodont dinosaur Bugenasaura infernalis (Ornithischia: Ornithopoda) from the Upper Cretaceous of North America. - Revue Pal�obiologie, Gen�ve 18 (2): 517–534. - Peter M. Galton - 1999. - Technical comment: dinosaur with a heart of stone. - Science 291 (5505): 783a. - Timothy R. Rowe, Earle F. McBride & Paul C. Sereno - 2001. - Reply: dinosaur with a heart of stone. - Science 291 (5505): 783a. - Dale A. Russel, Paul E. Fisher, Reese F. Barrick & Michael K. Stoskopf - 2001. - Taxonomic revision of the basal neornithischian taxa Thescelosaurus and Bugenasaura. - Journal of Vertebrate Paleontology 29 (3): 758–770. - Clint A. Boyd, Caleb M. Brown, Rodney D. Scheetz & Julia A. Clarke - 2009. - Histological, chemical, and morphological reexamination of the ‘heart’ of a small Late Cretaceous Thescelosaurus. - Naturwissenschaften 98 (3): 203–211. - Timothy P. Cleland, Michael K. Stoskopf & Mary H. Schweitzer - 2011. - A new basal ornithopod dinosaur (Frenchman Formation, Saskatchewan, Canada), and implications for late Maastrichtian ornithischian diversity in North America. - Zoological Journal of the Linnean Society 163 (4): 1157–1198. - Caleb M, Brown, Clint A. Boyd & Anthony P. Russel - 2011.








